Le déclin cognitif est un aspect inévitable du vieillissement qui affecte d’importantes compétences comportementales et cognitives. Les programmes de formation peuvent améliorer la cognition, mais il manque une caractérisation précise des fondements psychologiques et neuronaux qui soutiennent les différents programmes de formation. Ici, nous avons évalué l'effet et le maintien (suivi de 3 mois) de programmes de formation en musique et en arts visuels de 3 mois sur l'activité cérébrale neuroélectrique chez les personnes âgées en utilisant un plan d'intervention partiellement randomisé. Au cours des séances de tests pré, post et de suivi, les participants ont effectué une brève évaluation neuropsychologique. L'EEG à haute densité a été mesuré tandis que les participants se voyaient présenter des paradigmes auditifs étranges (tonalités de piano, voyelles) et lors d'une tâche visuelle GoNoGo. Aucun des deux programmes de formation n’a eu d’impact significatif sur les mesures psychométriques, par rapport à un groupe témoin non actif. Cependant, les participants inscrits aux programmes de formation en musique et en arts visuels ont montré une amélioration des réponses auditives évoquées aux sons du piano qui ont persisté jusqu'à 3 mois après la fin de la formation, suggérant des effets neuroplastiques robustes et durables. La formation en musique et en arts visuels a également modulé le traitement visuel au cours de la tâche GoNoGo, bien que ces effets de formation aient été relativement de courte durée et aient disparu au bout de 3 mois de suivi. Notamment, les participants inscrits à la formation en arts visuels ont montré des changements plus importants dans la distribution de l’amplitude de la réponse visuelle évoquée (c’est-à-dire l’onde N1) que ceux du groupe musique ou témoin. À l’inverse, ceux inscrits dans le groupe musique ont montré une plus grande réponse associée au contrôle inhibiteur sur les zones frontales droites du cuir chevelu que ceux du groupe art visuel. Nos résultats révèlent une relation causale entre la formation artistique (musique et arts visuels) et les changements neuroplastiques dans les systèmes sensoriels, certains changements neuroplastiques étant spécifiques au programme d'entraînement.
Introduction
Le vieillissement est associé à des changements structurels et fonctionnels dans les régions temporales préfrontales, pariétales et médiales, qui ont été associés à des réponses sensorielles évoquées améliorées dans les zones auditives et visuelles ainsi qu'à des déficits d'attention, de mémoire et de fonctions exécutives (Dempster, 1992; Alain et Woods, 1999; West et Alain, 2000; Bugaiska et al., 2007; Kropotov et al., 2016; Aljondi et al., 2018). Certaines théories mettent l'accent sur le déclin des processus inhibiteurs lié à l'âge (Hasher et Zacks, 1988; Knight, 1994; West, 1996), qui impliquent la capacité de réguler les entrées sensorielles et le comportement entrants afin afin de contourner les prédispositions internes ou les leurres externes pour accomplir des actions ou des comportements plus appropriés (Diamond, 2013). Déclins liés à l'âge dans la régulation attentionnelle et le contrôle inhibiteur (May et al., 1999; Erber, 2013) entraînent des conséquences négatives pour les compétences perceptuelles et cognitives (Gazzaley et al., 2007; ;Lustig et al., 2007), ce qui peut se refléter dans la réactivité globale (amplitude) des réponses sensorielles évoquées (Bidelman et al., 2017). Les déclins du contrôle inhibiteur liés à l'âge peuvent également être associés à des troubles de la compréhension du langage (Pichora -Fuller et al., 1995, 2017) et peut conduire à une augmentation des comportements socialement inappropriés. Compte tenu de l'expansion rapide de la population vieillissante dans la société moderne, il est de plus en plus nécessaire d'identifier et de développer des interventions efficaces qui réduisent, stoppent, voire inversent le déclin du contrôle inhibiteur afin de préserver les capacités cognitives quotidiennes des personnes âgées.
Des études récentes ont montré qu'un engagement tout au long de la vie dans des activités musicales pourrait aider à maintenir le cerveau dans un état plus jeune (Rogenmoser et al., 2018), lents déclins cognitifs (Hanna-Pladdy et Gajewski, 2012; Parbery -Clark et al., 2012; Zendel et Alain, 2012; Bidelman et Alain, 2015) , et préserver, voire améliorer le fonctionnement neuronal chez les personnes âgées (Parbery-Clark et al. , 2011; Bidelman et al., 2014; Zendel et Alain, 2014). Par exemple, les musiciens plus âgés affichent des performances améliorées par rapport aux non-musiciens dans plusieurs domaines cognitifs, notamment la mémoire spatiale, la vitesse de traitement et la flexibilité cognitive (Parbery-Clark et al., 2011), ainsi que le contrôle inhibiteur (Moreno et al., 2014; Moussard et al., 2016). Une relation positive entre les années d'engagement musical et les performances cognitives impliquait en outre des changements dépendants de l'expérience dans les compétences comportementales des personnes âgées (Parbery-Clark et al., 2011, 2012; Bidelman et Alain, 2015).
Plusieurs études sur le développement ont été menées pour étudier l'effet de l'apprentissage du chant ou de la lecture d'un instrument de musique sur une période de temps. En utilisant des conceptions d'experts et d'intervention, les résultats ont montré des bénéfices sur différents types de processus cognitifs tels que le traitement verbal (Moreno et al., 2009; Seither- Preisler et al., 2014), renseignement (Schellenberg, 2004), lecture (Moreno et al., 2011) , traitement inhibiteur (Moreno et al., 2011), traitement auditif (Moreno et Farzan, 2015; Tierney et Kraus, 2015) et sur le développement général du cerveau (Hyde et al., 2009). Cependant, les données probantes démontrant jusqu’à présent les bienfaits de la formation musicale chez les personnes âgées sont souvent corrélationnelles et non causales. Très peu d’études ont examiné, dans le cadre de conceptions longitudinales, si l’engagement dans une activité musicale pouvait améliorer les fonctions cognitives. Bien que des preuves suggèrent qu'une formation musicale à court terme (par exemple, des cours de piano) peut améliorer le fonctionnement exécutif et la mémoire de travail (Bugos et al., 2007; Seinfeld et al., 2013), les études antérieures souffrent souvent de limites méthodologiques (par exemple, absence de groupe témoin actif, manque de randomisation pour l'affectation des groupes) et n'ont pas mesuré les effets de l'intervention sur l'activité cérébrale. Ainsi, il reste à déterminer si la participation à des activités musicales entraînerait des changements neuroplasiques dans l’activité cérébrale et si ces changements pourraient être durables. Examiner l'activité neuronale avant et après un entraînement musical à court terme et la comparer à celle d'une autre activité artistique permettrait de mieux comprendre la plasticité cérébrale et les mécanismes de transfert potentiels dans le cerveau âgé.
L'art visuel est une forme de formation exigeante sur le plan cognitif qui pourrait être considérée comme aussi engageante que l'activité musicale. Les données probantes issues d'études de neuroimagerie suggèrent que l'activité est suscitée lors de l'engagement dans les arts visuels, y compris l'apprentissage d'objets visuels (Op de Beeck et Baker, 2010) et les activités motrices liées à la main (Draganski et al., 2004), induit des modifications neuroplastiques dans les zones cérébrales associées aux capacités de raisonnement spatial (Pollmann et von Cramon, 2000). La formation en arts visuels a également été associée à des différences structurelles dans les zones du cerveau liées au contrôle de la motricité fine et à la mémoire procédurale entre artistes et non-artistes (Chamberlain et al., 2014). Il y a donc des raisons de croire que la formation en arts visuels, tout comme la formation musicale, peut favoriser des améliorations cognitives chez les personnes âgées. Il est important de noter que la formation en arts visuels fournit un moyen d'évaluer si les changements neuroplastiques associés à la formation musicale sont spécifiques à la formation musicale elle-même ou s'ils reflètent des effets non spécifiques à la formation associés à l'engagement dans des programmes artistiques en général.
Dans cette étude, nous avons étudié l'impact à court terme (3 mois) de deux formes de formation engageante – l'enseignement de la musique et des arts visuels – sur les fonctions perceptuelles et cognitives des personnes âgées en utilisant une conception d'intervention en trois groupes (musique, art visuel et un contrôle sans contact). Nous avons également mesuré le maintien des effets avec un test de suivi 3 mois après l'arrêt de l'entraînement. Tous les participants ont effectué une brève évaluation neuropsychologique pour garantir que nos groupes étaient comparables avant la formation. Nous avons utilisé la même batterie après la formation et lors du suivi pour déterminer si la formation en musique et en arts visuels avait un impact sur le fonctionnement cognitif, tel que mesuré par des tests neuropsychologiques. Nous prévoyions que les participants du groupe de musique montreraient un fonctionnement cognitif plus élevé que ceux des groupes témoins actifs (arts visuels) et passifs. Nous avions prévu que les participants du groupe de musique montreraient un fonctionnement cognitif plus élevé après l'entraînement que ceux du groupe témoin. Nous nous attendions également à des améliorations potentielles suite à une formation en arts visuels, à titre d’hypothèse exploratoire.
De plus, nous avons enregistré les potentiels neuroélectriques liés aux événements (ERP) à l'aide de paradigmes auditifs étranges et lors d'une tâche visuelle GoNoGo à chaque séance (avant, après et suivi), afin d'évaluer les changements neuroplastiques liés à l'entraînement dans le traitement sensoriel. et fonctions exécutives. Ces paradigmes ont été choisis parce que des études antérieures ont montré des changements neuroplasiques associés à l'entraînement musical sur ces tâches (par exemple,Shahin et al., 2003; Zendel et Alain, 2009; Moussard et al., 2016 ). Nous avons émis l'hypothèse que chaque forme de formation artistique aurait un impact sur le traitement sensoriel dans la modalité entraînée, parallèlement aux avantages liés à la formation rapportés chez les enfants (par exemple,Fujioka et al., 2006; Moreno et al., 2011, 2014). Autrement dit, l'entraînement musical devait avoir un impact sur les ondes N1 et P2 des ERP auditifs, comme cela a été constaté précédemment chez les populations plus jeunes (par exemple,Tremblay et al., 2001; Reinke et al., 2003; Alain et al., 2010). Nous avions également prévu que l'entraînement musical améliorerait la capacité des auditeurs à remarquer les changements dans les stimuli auditifs, indexés par le décalage négatif (MMN), qui a également été observé chez les jeunes adultes (Tervaniemi et al., 1997; Nikjeh et al., 2009). Nous avons également émis l'hypothèse que l'entraînement aux arts visuels aurait un impact plus important que l'entraînement musical sur les réponses visuelles évoquées sensorielles telles que les ondes N1 et P2 au niveau des sites pariétaux-occipitaux.
De plus, nous nous attendions à observer des améliorations neuroplastiques dans les mécanismes cérébraux favorisant le contrôle inhibiteur dans le groupe de musique par rapport au groupe d'arts visuels et aux contrôles sans formation du même âge. Suite à la formation musicale, des changements étaient attendus dans les complexes N2 et P3 des ERP lors de la tâche visuelle GoNoGo, comme cela avait été constaté précédemment chez les plus jeunes (Brydges et al., 2014; Moreno et al., 2014) et plus anciens (Moussard et al., 2016). Comme le montrent des études antérieures menées auprès de jeunes adultes, l'entraînement musical a induit une amélioration de l'activité de l'hémisphère droit ( Moreno et al., 2014). Ainsi, nous avons émis l’hypothèse que le groupe de musique présenterait une augmentation de l’amplitude de l’ERP dans l’hémisphère droit après l’entraînement. Des recherches antérieures ont également montré les avantages durables de la formation musicale qui persistent des années après la fin de la formation (White-Schwoch et al., 2013). Par conséquent, nous nous attendions à ce que les améliorations cérébrales et comportementales persistent après un suivi de 3 mois. Cette découverte établirait qu'une formation artistique relativement engageante pourrait produire des effets durables sur les fonctions neuronales et comportementales des personnes âgées bien après la fin de l'enseignement.
Matériels et méthodes
Participants
Soixante personnes âgées en bonne santé ayant une formation préalable limitée en musique ou en arts visuels ont été recrutées dans la région du Grand Toronto. Pour les phases pré et post-formation, cinq participants ont été perdus à cause de l'attrition et deux en raison de problèmes techniques lors de l'enregistrement EEG, ce qui donne 17 participants dans le groupe de musique (trois hommes), 19 dans le groupe d'arts visuels (deux hommes), et 17 dans le groupe témoin sans contact (trois hommes). Les groupes ne différaient pas en termes d'âge (p = 0,82 ; musique : M = 67,7, SD = 5,8 ans ; arts visuels : M = 68,9, SD = 6 ans ; = 68,5, SD = 6 ans), années d'éducation formelle (p = 0,55 ; musique : M = 16,4, SD = 2,6 ans ; arts visuels : M = 17,2, SD = 2,3 ans ; contrôle : M = 16,9, SD = 1,5 ans), ou intelligence sur l'échelle abrégée d'intelligence de Wechsler -Deuxième édition (WASI-II FSIQ4; p = 0,51 ; musique : M = 114,2, SD = 10,5 ; art visuel : M = 115,8, SD = 11 ; contrôle : M = 111,8, SD = 13,9). Les participants ont été dépistés pour l'amusie et d'autres déficits auditifs ou musicaux qui auraient pu interférer avec l'étude à l'aide du Musical Ear Test (MET, Wallentin et al., 2010). Les trois groupes ont montré des scores similaires lors de l'évaluation psychométrique de base (toutes les valeurs p > 0,1). Après 3 mois, 15 participants du groupe de musique et 14 participants du groupe d'art visuel sont revenus pour des tests de suivi. Ces deux sous-groupes sont restés similaires en termes d'âge (p = 0,57), d'éducation (p = 0,31) et avaient une intelligence comparable sur WASI-II FSIQ4 (p = 0,39) au pré-test. L'étude a reçu l'approbation du comité d'éthique de la recherche de Baycrest et tous les participants ont fourni leur consentement éclairé écrit.
Étudier le design
Cette étude longitudinale comprenait quatre phases : pré-test, formation de 3 mois, post-test et test de suivi de 3 mois (Figure 1). Au cours des 3 mois entre le post-test et le suivi, les participants ne se sont pas engagés dans des activités formelles de musique ou d'arts visuels. Lors des séances de pré-test, de post-test et de suivi, les participants ont été testés individuellement et étaient aveugles à nos hypothèses. Après le pré-test, les participants ont été affectés à une formation en musique ou en arts visuels de manière pseudo-aléatoire pour égaliser les différences préalables à la formation entre les groupes en termes de scores d'intelligence et de mesures démographiques de base (sexe, âge et années d'études). Un groupe témoin passif supplémentaire a ensuite été recruté afin de collecter des données permettant de distinguer les effets potentiels de l'entraînement des effets test-retest.
Figure 1

Figure 1. Schematic of the study design.
Programmes de formation
Les participants formés ont reçu un enseignement et des activités en classe en groupe dans le cadre de leur formation respective par un enseignant professionnel (c'est-à-dire deux enseignants : un professeur de musique et un professeur d'arts visuels) au Royal Conservatory of Music de Toronto pendant 3 mois (36 séances d'une heure, trois fois par semaine). Le groupe de musique était engagé dans la création musicale en utilisant des percussions corporelles, de la voix et des instruments de musique sans tonalité. Ils ont également appris le solfège musical de base ainsi que les concepts de mélodie et d’harmonie grâce au chant de canons simples. Le groupe d'art visuel a appris les techniques de base du dessin et de la peinture, analysé le travail d'artistes célèbres et créé des peintures originales de paysages, de natures mortes et d'autoportraits. Tout le matériel leur a été fourni (c'est-à-dire des instruments pour le cours de musique et tout le matériel pour dessiner et peindre dans le cours d'art visuel).
Procédure
Les tests (EEG et tests psychométriques) ont eu lieu en laboratoire sur deux jours différents et ont duré environ 1,5 à 2 heures par séance. Pour les tests EEG, les enregistrements ont eu lieu dans une pièce protégée acoustiquement et électriquement.
Évaluation psychométrique
Wechsler Abbreviated Scale of Intelligence-Deuxième édition (WASI-II)
WASI-II évalue l'intelligence à l'aide de quatre sous-tests différents, y compris des mesures de compréhension verbale : (1) vocabulaire et (2) similarités ; et mesures du raisonnement perceptuel : (3) Conception de blocs et (4) Raisonnement matriciel. Un score composite (FSIQ4) a ensuite été calculé pour refléter le QI global.
Durée des mots en avant et en arrière
La tâche Word Span mesurait la mémoire de travail verbale, avec une condition avant et une condition arrière. L'expérimentateur lit des séquences de mots avec une pause d'une seconde entre les mots. Les listes de mots étaient organisées en longueurs de séquence (spans) de deux à huit. Il y avait deux versions du test qui étaient contrebalancées au fil des sessions de test. Chaque séquence comportait deux essais. Les tests se sont poursuivis jusqu'à ce que l'expérimentateur ait enregistré tous les mots rappelés de chaque liste. Un score total d'étendue a été calculé pour chaque condition en tant que nombre total d'éléments correctement rappelés dans tous les essais (score maximal d'étendue de mots = 70).
Test de Stroop
Une version papier comportant trois sous-tests du test de Stroop a été utilisée pour mesurer l'inhibition cognitive et la vitesse de traitement (Stroop, 1935). Le sous-test 1 impliquait la lecture de mots en couleur imprimés en caractères noirs, tandis que le sous-test 2 consistait à nommer la couleur des carrés. Le sous-test 3 nécessitait de nommer la couleur dans laquelle le mot était imprimé alors que le nom de la couleur écrit différait (c'est-à-dire une condition incongrue). Le temps de réponse a été mesuré pour chaque sous-test. Le score d'interférence a été calculé en soustrayant le temps de réponse au sous-test 3 de celui du sous-test 2. L'effet d'interférence s'est également reflété dans le nombre d'erreurs au sous-test 3.
Test de vocabulaire en images Peabody informatisé (PPVT)
Le Peabody Picture Vocabulary Test (PPVT) a été utilisé pour mesurer le vocabulaire réceptif ( Dunn et Dunn, 2007). Dans ce test, les participants voyaient quatre images sur l’écran pendant que l’ordinateur prononçait un mot. Leur tâche consistait à cliquer sur l’image qui caractérisait le mieux le sens du mot. Nous avons utilisé le nombre de réponses correctes dans l'analyse.
Symbole numérique (sous-test du WAIS-R)
Les participants ont eu 2 minutes pour copier les symboles sous les nombres selon une clé de codage où un nombre correspondait à un symbole spécifique. Le test a été noté en fonction du nombre de symboles correctement complétés.
Enregistrement EEG et analyse des données
Pour sonder les changements liés à l'entraînement dans l'activité cérébrale soutenant les processus de contrôle perceptuel et exécutif, nous avons mesuré l'activité neuroélectrique (électroencéphalogramme, EEG) à l'aide de paradigmes auditifs étranges et au cours d'une tâche visuelle GoNoGo. Les mêmes stimuli ont été utilisés lors de chacune des trois séances de tests (pré, post et suivi), tandis que l'ordre des essais a été randomisé entre les participants et les séances.
Paradigmes auditifs bizarres
Le traitement auditif a été évalué via la négativité N1, P2 et les mésappariements (MMN) enregistrés avant et après l'entraînement à l'aide de deux séquences étranges consistant en un contraste sonore de musique ou de parole. Pour la condition musicale, deux sons de piano synthétisés, Eb4 (F0 = 314 Hz) et D4 (F0 = 294 Hz) ont servi respectivement de jetons standard et déviants. Les deux notes ne différaient que par leur hauteur et étaient par ailleurs identiques en termes de caractéristiques acoustiques, notamment l'amplitude et la durée globales (500 ms). Pour la condition de parole, deux voyelles françaises produites par une locutrice native, /u/ et /ou/, fonctionnaient respectivement comme stimulus standard et déviant. Les deux jetons vocaux avaient une durée similaire (280 ms), une fréquence fondamentale vocale moyenne (F0 : ∼240 Hz), une amplitude et des fréquences de premier/troisième formant ; seul leur deuxième formant différait pour donner les deux timbres de voyelles distincts (/u/ : ∼1 850 Hz ; /ou/ : ∼750 Hz). L'ordre de présentation était pseudo-aléatoire, de sorte qu'au moins un stimulus standard précédait un déviant. Les stimuli ont été présentés avec une asynchronie de début de stimulus de 750 ms et délivrés de manière binaurale via des écouteurs à insert ER-3 (Etymotic Research) à une intensité de niveau de pression acoustique de 80 dB. Un total de 510 normes et 90 déviants (soit un ratio de 85/15 %) ont été collectés pour les conditions de parole et de musique.
Visual GoNoGo task
Les participants se sont vu présenter un triangle ou des carrés géométriques blancs ou violets au centre de l'écran situé à environ 1 m du participant. Avant chaque essai, une croix de fixation blanche apparaissait sur fond noir pendant une durée variable (500 à 1 000 ms), puis une forme géométrique apparaissait au centre de l'écran pendant 500 ms. Les participants devaient appuyer sur le bouton droit de la souris en réponse aux formes blanches (probabilité de 80 %) et ne pas répondre aux formes violettes (probabilité de 20 %). L'expérience comprenait 200 essais (160 essais Go et 40 essais NoGo). Un bloc de pratique de 20 essais a été utilisé pour familiariser les participants avec la tâche. Au cours de la tâche, les participants n'ont reçu aucun retour sur leur performance. Les taux de précision ont été enregistrés pour les essais Go et NoGo, et les temps de réaction ont été enregistrés pour les essais Go.
EEG recording and data processing
L'EEG a été enregistré à partir de 66 électrodes du cuir chevelu à l'aide d'un système d'acquisition BioSemi Active Two (BioSemi V.O.F., Amsterdam, Pays-Bas). Le montage de l'électrode était conforme au capuchon d'électrode BioSemi basé sur le système 10/20 et comprenait une électrode active de détection de mode commun et une électrode passive de jambe droite entraînée servant de masse. Dix électrodes supplémentaires ont été placées sous la racine des cheveux (les deux mastoïdes, les deux points pré-auriculaires, le canthus externe de chaque œil, l'orbite inférieure de chaque œil, deux électrodes faciales) pour surveiller les mouvements oculaires et couvrir uniformément tout le cuir chevelu. Ce dernier est important car nous avons utilisé une référence moyenne (c’est-à-dire la moyenne de tous les canaux EEG du cuir chevelu comme référence pour chaque canal EEG) pour les analyses ERP. L'activité neuroélectrique a été numérisée en continu à une fréquence de 512 Hz avec une bande passante de DC-100 Hz et stockée pour une analyse hors ligne. Les analyses hors ligne ont été effectuées à l'aide du logiciel Brain Electrical Source Analysis (BESA, version 6.1 ; MEGIS GmbH, Gräfelfing, Allemagne).
Les EEG continus ont d’abord été filtrés numériquement avec des filtres passe-haut de 0,5 Hz (avant, 6 dB/octave) et passe-bas de 40 Hz (phase zéro, 24 dB/octave). Pour les potentiels évoqués auditifs, la période d'analyse comprenait 100 ms d'activité pré-stimulus et 500 ms d'activité post-stimulus synchronisées sur l'apparition du son. Pour le GoNoGo, l’époque d’analyse comprenait 200 ms d’activité pré-stimulus et 1 000 ms d’activité post-stimuli synchronisées avec l’apparition des stimuli visuels. Les segments EEG contaminés par des clignements et des saccades ont été corrigés à l'aide de BESA. Les clignements des yeux et les mouvements latéraux ont d'abord été identifiés dans l'EEG continu, puis modélisés à l'aide de la correction des artefacts avec un modèle de substitution. Après correction oculaire, les traces ont ensuite été analysées à la recherche d'artefacts et les époques, y compris les déviations dépassant 120 μV, ont été marquées et exclues de l'analyse. Les époques restantes ont été moyennées en fonction de la position de l'électrode, du type d'essai (par exemple, Go, NoGo) et de la session (c'est-à-dire pré-entraînement, post-entraînement, suivi). Chaque moyenne a été corrigée en fonction de la ligne de base par rapport à l'intervalle précédant le stimulus.
Pour les paradigmes bizarres, la proportion d'essais inclus dans les ERP auditifs variait de 283 à 510 essais pour les stimuli standards et de 48 à 98 pour les stimuli déviants. Le nombre d'essais était comparable pour les séances pré-(Standard : M = 487 ; Déviant : M = 85) et post-entraînement (Standard : M = 484 ; Déviant : M = 86), et était similaire pour les contrôles (Standard : M = 493 ; Déviant : M = 88), groupes de musique (Standard : M = 478 ; Déviant : M = 84) et d'arts visuels (Standard : M = 484 ; Déviant : M = 85). Pour la tâche GoNoGo, le nombre d'essais inclus dans les ERP visuels variait de 78 à 160 essais pour la condition Go et de 17 à 40 essais pour la condition NoGo. En ce qui concerne les paradigmes bizarres, le nombre d'essais était comparable avant (Go : M = 141 ; NoGo : M = 34) et après (Go : M = 144 ; NoGo : M = 35) l'entraînement, et était similaire pour le contrôle (Go : M = 148 ; NoGo : M = 36), de musique (Go : M = 141 ; NoGo : M = 33) et d'arts visuels (Go : M = 139 ; NoGo : M = 34).
Analyses statistiques
Les effets de l'entraînement sur les mesures psychométriques ont été évalués à l'aide d'ANOVA à mesures répétées. Nous nous sommes concentrés sur les interactions qui impliquaient le groupe et la session comme facteurs. Bien qu’ils ne soient pas au cœur de l’objectif de la présente étude, nous rapportons également d’autres effets et interactions principaux par souci d’exhaustivité. Pour les évaluations psychométriques, nous avons utilisé la méthode Benjamini-Hochberg pour ajuster la valeur p familiale pour plusieurs comparaisons (q = 0,1, m = 30 et p = 0,05), ce qui a donné une valeur p de 0,005 pour la signification.
Les analyses ERP se sont concentrées sur des fenêtres temporelles prédéfinies et des groupes d'électrodes motivés par des études antérieures (par exemple,Reinke et al., 2003; Moreno et al., 2014; Moussard et al. ., 2016). Pour les paradigmes étranges, les effets de l'entraînement sur les déflexions N1 et P2 ont été évalués pendant les intervalles de 90 à 130 ms et de 170 à 210 ms au niveau des sites fronto-centraux du cuir chevelu (F1, Fz, F2, FC1, FCz, FC2, C1, Cz ,C2). La détection automatique des changements a été quantifiée à l'aide des ondes de différence entre les ondes standard et déviantes. L'amplitude et la latence maximales ont été quantifiées au cours de la fenêtre de 100 à 250 ms en utilisant les mêmes groupes d'électrodes que les ondes N1 et P2. Pour la tâche GoNoGo, le traitement des stimuli go et nogo a été évalué pendant l'intervalle de 165 à 205 ms (c'est-à-dire N1) et l'intervalle de 210 à 280 ms au niveau des électrodes pariéto-occipitales de gauche (PO7, P5, P7) et de droite. hémisphère (PO8, P6, P8). Les effets de la musique et de l'entraînement artistique sur la capacité à inhiber une réponse motrice dans les essais nogo ont été quantifiés en comparant l'amplitude moyenne pour l'intervalle de 375 à 475 ms mesuré sur la gauche (F1, F3, AF3) et la droite (F2, F4, AF4) régions frontales du cuir chevelu. Ces groupes d'électrodes ont été choisis car il a été démontré que l'entraînement musical chez les jeunes adultes module l'amplitude visuelle de l'ERP GoNoGo sur l'hémisphère droit (Moreno et al., 2014). Ainsi, nous avions prévu que les activités musicales chez les personnes âgées montreraient également des changements dans l'amplitude de l'ERP dans l'hémisphère droit après l'entraînement.
Les effets de la formation artistique sur les réponses évoquées auditives (c'est-à-dire N1, P2, MMN) ont été évalués à l'aide d'une ANOVA à modèle mixte utilisant des groupes (contrôle, musique, arts visuels) comme facteur entre les sujets et le type de séquence (musique vs . parole) et le type de stimulus (standard, déviant) comme facteurs intra-sujets. Les effets de la formation artistique sur les réponses visuelles évoquées enregistrées au cours de la tâche GoNoGo ont été évalués à l'aide d'une ANOVA à modèle mixte avec le groupe (contrôle, musique, art visuel) comme facteurs inter-sujets, le type de stimulus (essais Go, NoGo) et l'hémisphère ( droite, gauche) comme facteurs intra-sujets. Nous avons utilisé la méthode Benjamini-Hochberg pour ajuster la valeur p familiale pour des comparaisons multiples avec q = 0,1, m = 160 (c'est-à-dire le nombre de p-valeurs) et p = 0,05 (Hochberg et Benjamini, 1990), ce qui donne une valeur p de 0,025 pour la signification.
Résultats
Évaluation psychométrique : effets de l'entraînement
Les analyses des performances au test neuropsychologique ont révélé une interaction groupe × session sur le sous-test Stroop 2 [vitesse de dénomination des couleurs ; F(2,50) = 6,16, p = 0,004, η2 = 0,20], où le groupe de musique a amélioré sa vitesse de dénomination entre la pré- et la post-évaluation [t(16 ) = 3,32, p = 0,004, Figure 2]. Il y avait également un effet principal de la session pour la tâche Symbole numérique, avec de meilleures performances au post-test [par rapport au pré-test; F(1,50) = 15,98, p < 0,000, η2 = 0,24], mais aucune interaction avec le groupe. Il n’y avait pas d’autres différences significatives de performance entre les séances avant et après l’entraînement. Dans le groupe musique, une analyse de suivi a montré que le gain en vitesse de réponse était maintenu 3 mois après l'entraînement (contraste post-test vs suivi, p > 0,1) .
Figure 2
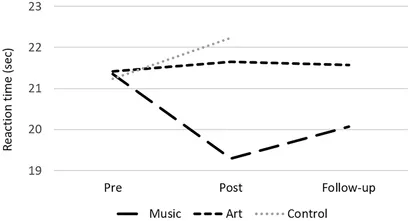
Figure 2. Temps de réaction moyen au sous-test Stroop 2 (nommage des couleurs) pour les groupes musique, art et contrôle au pré-test, au post-test et au suivi.
Effets de la formation en musique et en arts visuels sur le traitement auditif
Figures 3, 4 afficher les ERP auditifs moyens du groupe provoqués par des stimuli standards et déviants ainsi que la différence correspondante ondes dans les deux séquences (tonalités de piano et voyelles) lors des séances pré et post-formation, ainsi que du sous-ensemble de participants qui ont effectué le suivi. Les ERP enregistrés sur le cuir chevelu comprenaient des déviations prototypiques à ∼60, ∼110 et ∼195 ms après l'apparition du son (c'est-à-dire P1-N1-P2). Le MMN a été isolé comme la différence d’activité neuronale entre les stimuli standards et déviants (étranges). Un participant du groupe de musique et deux du groupe d'art ont été exclus des analyses car les données de l'une des deux séquences n'étaient pas disponibles en raison de problèmes techniques.
Figure 3

Figure 3. Groupez les potentiels liés aux événements moyens provoqués par les sons de piano standards et déviants ainsi que l’onde de différence correspondante (c’est-à-dire MMN). Ici et partout, la négativité est tracée vers le haut. Les formes d’onde proviennent de l’électrode fronto-centrale médiane (FCz).
Figure 4

Figure 4. Groupez les potentiels liés à l’événement moyen provoqués par des stimuli de voyelles standard et déviants ainsi que l’onde de différence correspondante (c’est-à-dire MMN). Les formes d’onde proviennent de l’électrode fronto-centrale médiane (FCz).
Training Effects on Auditory ERPs
Tout d'abord, nous avons examiné si la formation en musique et en arts visuels modulait le traitement cortical précoce indexé par les ondes N1 et P2. L'ANOVA sur l'amplitude moyenne N1 (90–130 ms) a donné une interaction groupe × session × type de séquence significative [F(2,47) = 4,30,p = 0,019, η2 = 0,16]. Pour mieux comprendre cette interaction à trois voies, nous avons examiné les effets de l'entraînement en fonction du type de séquence. Pour les sons de piano, les groupes de musique et d'art ont montré une amplitude N1 plus grande au post-test qu'au pré-test (p < 0,005 dans les deux cas). Pour le groupe témoin, il n’y avait aucune différence d’amplitude N1 entre le pré-test et le post-test. Pour les stimuli vocaliques, il n'y avait pas de différence significative dans l'amplitude N1 entre le pré-test et le post-test (p > 0,19 dans les trois groupes). Ainsi, alors que l'amplitude N1 provoquée par les stimuli vocaliques était peu affectée par l'entraînement, le N1 généré par les sons du piano était plus grand au post-test qu'au pré-test dans les groupes de formation en musique et en arts visuels (Figures 3, 4). L'interaction entre le groupe × la séance × le type de stimulus n'était pas significative.
Nous avons également observé un effet principal de session [F(1,47) = 10,15, p = 0,003, η2 = 0,18], et une interaction de type session × séquence [F(1,47) = 6,06, p = 018, η2 = 0,11], avec une augmentation plus importante de l'amplitude N1 au post-test pour le son du piano que pour les stimuli vocaliques. L'effet principal du type de stimulus était significatif [F(1,47) = 52,19, p < 0,001, η2 = 0,53], reflétant une amplitude N1 plus grande pour les stimuli déviants que standards. L'interaction entre la séance et le type de stimulus était également significative [F(1,47) = 12,82, p = 0,001, η2 = 0,21], ce qui était dû à une plus grande différence entre standard et déviant au post-test qu'au pré-test.
Pour l'amplitude moyenne P2 (175-215 ms), le groupe × séance n'était pas significatif, pas plus que le type de stimulus groupe × séance ×. Cependant, l'interaction de type groupe × session × séquence s'est approchée du niveau de signification corrigé [F(2,47) = 3,63, p = 0,034, η2 = 0,13]. Pour les sons de piano, les groupes de musique et d'arts visuels ont montré une amplitude P2 plus grande au post-test qu'au pré-test (p < 0,005 dans les deux cas). Pour le groupe témoin, il n’y avait aucune différence d’amplitude P2 entre le pré-test et le post-test. Pour les stimuli vocaliques, il n'y avait pas de différence significative dans l'amplitude P2 entre les séances (p > 0,65 dans les trois groupes). Quant à l'amplitude N1, l'onde P2 provoquée par les stimuli vocaliques était peu affectée par l'entraînement tandis que l'onde P2 générée par les sons du piano était plus grande au post-test qu'au pré-test dans les groupes de formation en musique et en arts visuels (Figures 3, 4).
Figures 3, 4 montrer le MMN moyen entre les groupes. Pour la latence MMN, l'interaction groupe × session n'était pas significative [F(2,47) = 1,70, p = 0,193, η2 = 0,07], ni l'interaction de type groupe × séquence (F < 1). L'interaction à trois groupe × session × type de séquence n'était pas significative [F(2,47) = 1,80, p = 0,177, η 2 = 0,071]. Cependant, l'ANOVA a produit un effet principal de type séquence [F(1,47) = 49,06, p < 0,001, η2 = 0,51], avec le MMN provoqué par les sons du piano (M = 150 ms, SE ;= 2,5 ms) avec un pic plus tôt que les voyelles suscitées par le MMN (M = 182 ms, SE = 4,1 ms). L'effet principal de la séance était également significatif [F(1,47) = 16,42, p < 0,001, η2 = 0,23], avec un pic de latence plus précoce au post-test qu'au pré-test.
Effets de l'entraînement sur le MMN
Figures 3, 4 montrer le MMN moyen entre les groupes. Pour la latence MMN, l'interaction groupe × session n'était pas significative [F(2,47) = 1,70, p = 0,193, η2 = 0,07], ni l'interaction de type groupe × séquence (F < 1). L'interaction à trois groupe × session × type de séquence n'était pas significative [F(2,47) = 1,80, p = 0,177, η 2 = 0,071]. Cependant, l'ANOVA a produit un effet principal de type séquence [F(1,47) = 49,06, p < 0,001, η2 = 0,51], avec le MMN provoqué par les sons du piano (M = 150 ms, SE ;= 2,5 ms) avec un pic plus tôt que les voyelles suscitées par le MMN (M = 182 ms, SE = 4,1 ms). L'effet principal de la séance était également significatif [F(1,47) = 16,42, p < 0,001, η2 = 0,23], avec un pic de latence plus précoce au post-test qu'au pré-test.
For the MMN peak amplitude, the group × session interaction was not significant [F(2,47) = 2.55, p = 0.089, η2 = 0.10], nor was the group × sequence type (F < 1). However, the MMN was larger for vowel than piano tones [F(1,47) = 49.22, p < 0.001, η2 = 0.51]. All other main effects and interactions were not significant. Thus, neither music nor visual art training had a significant effect on the automatic change detection process as measured by the MMN.
Rétention de suivi
Trois mois après la fin de la formation, un sous-groupe de participants (musique N = 15, arts visuels N = 14) est revenu pour une évaluation de suivi et une évaluation des effets de rétention à long terme. Un participant de chaque groupe a été exclu de l'analyse car il lui manquait l'une des conditions expérimentales en raison de problèmes techniques. L'échantillon restant comprenait 14 personnes du groupe de musique et 13 personnes du groupe d'arts visuels.
L'ANOVA sur l'amplitude moyenne N1 avec la session (pré, post, suivi) comme facteur inter-sujet et la séquence (tonalités de piano, voyelles) et le type de stimulus (standard, déviant) comme facteurs intra-sujet, a donné un interaction significative entre session × type de séquence [F(2,50) = 4,47, p = 0,016, η2 = 0,15]. Pour mieux comprendre cette interaction, nous avons effectué des ANOVA distinctes pour les stimuli du piano et des voyelles. Pour les sons de piano, l'ANOVA a révélé un effet principal de session [F(2,50) = 11,33, p < 0,001, η2 = 0,31]. Les comparaisons par paires ont révélé une augmentation significative de l'amplitude N1 au post-test et au suivi par rapport au pré-test (p < 0,001 dans les deux cas). Il n'y avait aucune différence d'amplitude N1 entre le post-test et le suivi (p = 0,144). L'interaction groupe × séance n'était pas significative (F < 1), ni le type de stimulus groupe × séance × [F(2,50) = 1,67, p = 0,192, η2 = 0,31]. Pour les stimuli vocaliques, l'effet principal de la séance n'était pas significatif, pas plus que l'interaction groupe × séance (F < 1 dans les deux cas). Quant au N1, l'ANOVA sur l'amplitude moyenne P2 a donné une interaction session × séquence [F(2,50) = 6,27, p = 0,004, η2 = 0,20]. Pour les sons de piano, l'effet principal de la séance était significatif [F(2,50) = 10,29, p < 0,001, η2 = 0,29]. Les comparaisons par paires ont révélé une amplitude P2 plus grande au post-test et au suivi qu'au pré-test (p < 0,001 dans les deux cas). Il n'y avait aucune différence d'amplitude P2 entre le post-test et le suivi (p = 0,51). Pour les stimuli vocaliques, l'effet principal de la session n'était pas significatif (F < 1), pas plus que l'interaction groupe × session (F < 1).
En résumé, la formation en musique et en arts visuels a modulé le traitement auditif, qui a été conservé 3 mois après la fin de la formation.
Effets de la formation en musique et en arts visuels sur les ERP pendant la tâche Visual Go NoGo
Données comportementales
Les trois groupes ont affiché des performances maximales dans les essais Go, avec peu (voire aucun) de faux positifs dans les essais NoGo (voir Tableau 1). Il n’y avait aucune différence entre les groupes ni entre les séances pré- et post-test. L'interaction groupe × session n'était pas significative pour les mesures de précision ou de temps de réponse.
Table 1

Table 1. Résultats comportementaux sur la tâche GoNoGo.
Effets de la formation sur les ERP visuels
Figure 5 montre les ERP visuels moyens de groupe obtenus au cours Essais Go et NoGo ainsi que les ondes différentielles correspondantes au niveau des sites pariétaux-occipitaux. Les ERP visuels enregistrés sur le cuir chevelu comprenaient des déviations prototypiques à environ 100, environ 185 et environ 325 ms après le début du stimulus (c'est-à-dire P1-N1-P2). L'onde de différence entre les essais Go et NoGo révèle un traitement associé à la suppression de la réponse motrice.
Figure 5
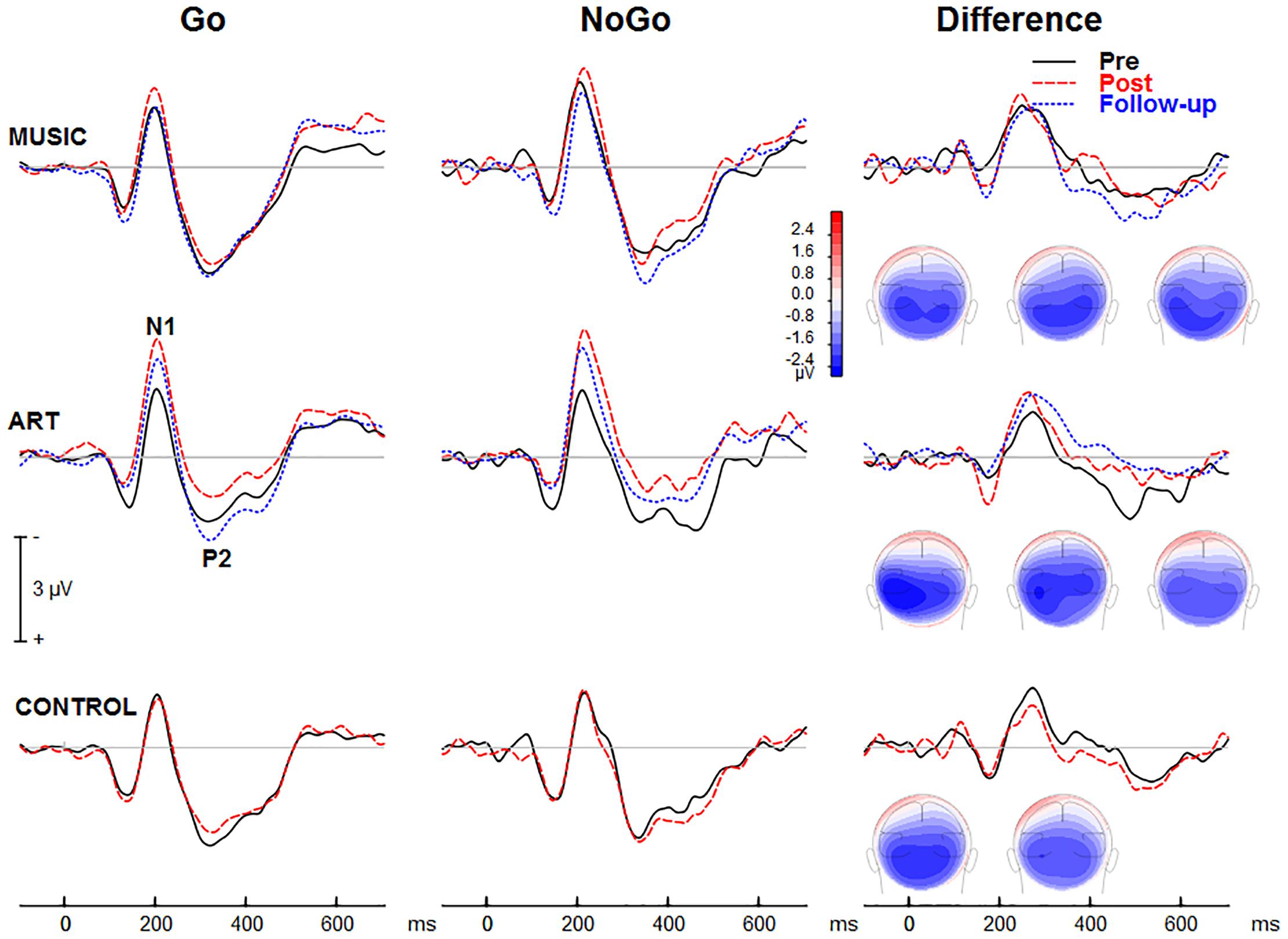
Figure 5. Groupez les potentiels liés à l’événement moyen suscités lors de la tâche visuelle GoNoGo sur la zone du cuir chevelu pariétal droit (c’est-à-dire l’électrode P6). Les cartes iso-contours (vue de dos) montrent la distribution d'amplitude pour la tension moyenne entre 210 et 280 ms. De gauche à droite, les cartes montrent respectivement la distribution au pré-test, au post-test et au suivi.
Nous avons d'abord examiné si la formation en musique et en arts visuels modulait le traitement cortical visuel précoce indexé par les ondes N1 et P2. L'ANOVA sur l'amplitude moyenne N1 (165–205 ms) a donné un groupe × séance × hémisphère significatif [F(2,50) = 4,00, p ;= 0,024, η2 = 0,14]. Pour mieux comprendre cette interaction à trois voies, nous avons examiné les effets de l'entraînement en fonction de l'hémisphère. Pour les zones pariétales-occipitales gauches, le groupe musique et arts visuels a montré une amplitude visuelle N1 comparable avant et après l'entraînement (p > 0,5 dans les deux cas) alors que la N1 dans le groupe témoin était plus grande. au post-test qu'au pré-test (p < 0,01). Pour les zones pariétales-occipitales droites, le groupe musique et le groupe témoin ont montré une amplitude N1 comparable au pré-test et au post-test, tandis que l'amplitude N1 était significativement améliorée dans le groupe art visuel (p = 0,003). Alors que l'amplitude N1 mesurée sur l'hémisphère gauche était peu affectée par l'entraînement, l'amplitude N1 enregistrée sur l'hémisphère droit était plus grande au post-test qu'au pré-test dans le groupe des arts visuels uniquement. Aucune autre interaction impliquant un groupe et des sessions n'était significative.
L'ANOVA omnibus a également montré que l'onde N1 était plus grande au post-test qu'au pré-test [F(1,50) = 6,21, p ;= 0,016, η2 = 0,11]. L'effet principal du type de stimulus était également significatif [F(1,50) = 18,78, p < 0,001, η2 = 0,27], avec une amplitude N1 plus grande pendant les essais NoGo que Go. Cette dernière pourrait refléter une négativité liée à l'attention qui chevauche l'onde N1 plutôt qu'une modulation de la réponse N1.
Pour l'amplitude visuelle moyenne P2 (305–345 ms), il n'y a pas eu d'interaction significative entre le groupe et la séance, et l'interaction groupe × séance × hémisphère n'a pas non plus été significative [F(2,50) = 1,35, p = 0,269, η2 = 0,05]. L'effet principal de la séance n'était pas significatif [F(1,50) = 2,23, p = 0,142, η2 ;= 0,04]. Cependant, l'interaction session × hémisphère était significative [F(1,50) = 4,14, p = 0,047, η2 = 0,08]. Alors que la P2 mesurée sur l'hémisphère gauche était comparable entre les séances pré et post-entraînement, la P2 enregistrée sur la zone pariéto-occipitale droite était plus grande au post-test.
Effets liés à NoGo
Les ERP visuels obtenus lors des essais NoGo ont montré un déplacement négatif, qui pourrait être expliqué par des effets liés à l'attention superposés sur le N1 et le P2. Ceci est mieux illustré par la vague de différence entre les essais Go et NoGo. Cette onde de différence a révélé une composante négative au niveau des sites pariétaux-occipitaux qui culmine environ 230 à 240 ms après le début du stimulus. L'amplitude moyenne de 210 à 280 a été utilisée pour les hémisphères gauche (PO7, P5, P7) et droit (PO8, P6, P8).
L'effet principal du groupe n'était pas significatif [F(2,50) = 1,41, p = 0,253, η2 = 0,05], ni l'interaction groupe × session (F < 1). Cependant, il y avait une interaction groupe × séance × hémisphère significative [F(2,50) = 6,17, p = 0,004, η2 = 0,19]. Cela était dû à un changement dans la latéralité de la réponse entre la première et la deuxième séance dans le groupe d'art visuel (Figure 6). Autrement dit, au pré-test, l'amplitude de l'ERP était plus grande sur les sites pariétaux-occipitaux gauches, alors qu'après une formation en arts visuels, les ERP présentaient une distribution d'amplitude plus symétrique sur les hémisphères gauche et droit. Dans le groupe de contrôle et de musique, l'amplitude ERP était plus grande sur les hémisphères gauche et droit pendant les séances pré et post-entraînement.
Figure 6
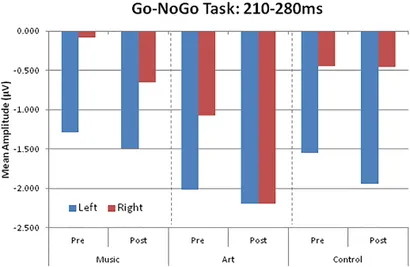
Figure 6. Groupez les amplitudes potentielles moyennes liées aux événements visuels sur les zones pariéto-occipitales gauche (PO7, P5, P7) et droite (PO8, P6, P8).
Il y a eu un effet principal de la session [F(1,50) = 6,21, p = 0,016, η2 = 0,11], avec une plus grande amplitude au post-test. Le principal effet de la condition était également significatif [F(1,50) = 124,47, p < 0,001, η2 = 0,71], ce qui reflète une négativité accrue générée par les essais NoGo par rapport aux essais Go.
Rétention de suivi
Nous avons examiné si les changements de latéralité persistaient 3 mois après la fin de la formation. Une ANOVA comparant les ERP enregistrés au cours des trois sessions du groupe artistique a révélé une interaction session × hémisphère [F(2,26) = 6,02, p = 0,013 ]. Des ANOVA séparées pour les hémisphères gauche et droit ont donné un effet principal de la séance uniquement pour les ERP enregistrés sur la zone pariétale-occipitale droite [hémisphère droit : F(2,26) = 4,35, < em>p = 0,029 ; hémisphère gauche : F < 1]. Les comparaisons par paires ont révélé une négativité accrue au post-test par rapport au pré-test et au suivi (p < 0,05 dans les deux cas). Les ERP au suivi ne différaient pas de ceux obtenus au pré-test (p = 0,693). Comme lors du pré-test, les ERP enregistrés lors du suivi étaient plus grands sur l’hémisphère gauche que sur l’hémisphère droit. En résumé, la formation artistique était associée à une augmentation des effets précoces liés à l'attention sur la zone pariétale-occipitale droite, qui n'était pas conservée 3 mois après la formation.
ERP associés à l'inhibition de la réponse
Les essais NoGo ont généré une modulation qui a culminé environ 425 ms après le début du stimulus sur les sites fronto-centraux (Figure 7). L'interaction groupe × séance × condition × hémisphère était significative [F(2,50) = 4,24, p = 0,020, η2 = 0,15]. Pour mieux comprendre cette interaction à quatre, nous avons effectué des ANOVA distinctes pour chaque groupe. Dans le groupe musique, l'effet principal de la condition était significatif [F(1,16) = 8,19, p = 0,011, η2 = 0,339], tout comme l'interaction session × condition [F(1,16) = 8,67, p = 0,010, η< sup>2 = 0,351], avec une augmentation plus importante de l'amplitude ERP au post-test pour la condition NoGo que Go. La condition × hémisphère était significative [F(1,16) = 13,31, p = 0,002, η2 = 0,454], reflétant une plus grande différence entre les essais Go et NoGo sur l'hémisphère gauche. La séance × hémisphère n'était pas significative (F < 1). Dans le groupe des arts visuels, le principal effet de la condition était significatif [F(1,18) = 21,18, p < 0,001, η2 = 0,541]. La condition session × était également significative [F(1,18) = 8,038, p = 0,011, η2 = 0,309], avec une augmentation plus importante de l'amplitude ERP au post-test pour la condition NoGo que Go. La session × hémisphère tend vers une signification [F(1,18) = 3,51, p = 0,077, η2 = 0,163]. Cependant, la séance × condition × hémisphère était significative [F(1,18) = 5,78, p = 0,027, η2 = 0,243]. Cela était dû à une plus grande différence d’amplitude ERP entre les essais Go et NoGo sur l’hémisphère gauche après l’entraînement. Dans le groupe témoin, le principal effet de la condition était significatif [F(1,16) = 17,12, p = 0,001, η = 0,517]. Cependant, ni l'effet principal de la session ni l'interaction session × condition n'étaient significatifs (F < 1 dans les deux cas). La condition × hémisphère était significative [F(1,16) = 29,04, p < 0,001, η2 = 0,645], reflétant une plus grande différence entre les essais Go et NoGo sur l'hémisphère gauche. Les autres effets ou interactions principaux n'étaient pas significatifs.
Figure 7
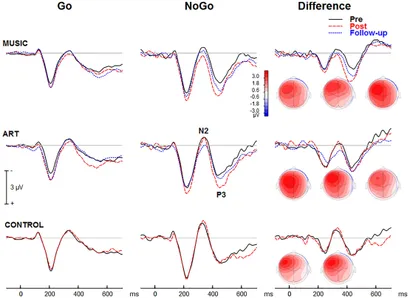
Figure 7. Potentiel moyen lié à un événement visuel provoqué lors de la tâche visuelle GoNoGo sur la région frontale droite du cuir chevelu (c'est-à-dire l'électrode F2). Les cartes iso-contours (vue du dessus) montrent la distribution d'amplitude pour la tension moyenne entre 375 et 475 ms. De gauche à droite, les cartes montrent respectivement la distribution au pré-test, au post-test et au suivi.
L'ANOVA sur l'amplitude moyenne (375–475 ms) a donné un effet principal de la séance [F(1,50) = 4,37, p = 0,042, η2 = 0,08], et un effet principal de la condition [F(1,50) = 43,59, p ≪ 0,001, η2 = 0,466]. Autrement dit, l'amplitude ERP était plus positive au post-test qu'au pré-test, et plus positive pour les essais NoGo que pour les essais Go. L'interaction session × condition était significative [F(1,50) = 12,50, p = 0,001, η2 = 0,20]. Cela reflète une plus grande différence entre les essais Go et NoGo (effet NoGo ERP) au post-test qu'au pré-test. L'interaction condition × hémisphère était également significative [F(1,50) = 62,18, p < 0,001, η2 = 0,55]. Cela était dû à des réponses NoGo plus importantes dans la région frontale gauche du cuir chevelu par rapport à la région frontale droite du cuir chevelu.
Rétention de suivi
Dans les groupes de musique et d’arts visuels, l’effet NoGo ERP était légèrement réduit lors du suivi par rapport au post-test, mais la différence n’était pas statistiquement significative. De plus, l’effet NoGo ERP lors du suivi ne différait pas des effets observés lors du pré-test. Ces résultats suggèrent que certains effets de l’entraînement persistaient trois mois après l’arrêt de l’entraînement. Cependant, ce résultat doit être interprété avec prudence en raison de la petite taille de l'échantillon au moment du suivi et de l'absence de différence entre le pré-test et la séance de suivi.
Discussion
Utilisant un plan d’intervention partiellement randomisé, notre étude révèle des changements dans les réponses cérébrales des personnes âgées suite à une formation artistique à court terme (engagement dans la musique et les arts visuels). Ainsi, nous fournissons de nouvelles preuves importantes suggérant une relation causale entre ces activités et les changements neuroplasiques chez les personnes âgées. Dans la présente étude, nous avons également observé certains effets spécifiques à l’entraînement en utilisant les paradigmes auditifs bizarres et visuels GoNoGo.
Formation artistique et traitement auditif
Dans les deux groupes, nous avons observé une augmentation de l'amplitude N1 et P2 après l'entraînement, qui a été conservée au moins 3 mois après la fin de l'entraînement. Bien qu'une augmentation de l'amplitude N1 et P2 ait été observée après l'entraînement dans plusieurs études (par exemple,Tremblay et coll., 2001; Reinke et coll. , 2003), il s'agit, à notre connaissance, de la première étude montrant une augmentation liée à l'entraînement de l'amplitude N1 et P2 à des stimuli non entraînés chez les personnes âgées. Dans la présente étude, l’effet de l’entraînement était plus prononcé pour les sonorités du piano. Cette différence d’effet N1 lié à l’entraînement entre les sons du piano et les voyelles pourrait être liée à la familiarité avec le matériel avant l’étude. Des recherches antérieures ont montré une augmentation de l'amplitude N1 et P2 avec une familiarité accrue (Sheehan et al., 2005; Alain et al., 2015 ). Pour la plupart des participants, les sons de la parole tels que les voyelles sont des stimuli surappris et très familiers, laissant moins de possibilités d'observer les effets neuroplastiques. En comparaison, les sons de piano utilisés dans la présente étude étaient plus nouveaux et bénéficiaient ainsi davantage de la formation. Le groupe de musique a peut-être ainsi bénéficié d’une exposition aux sons musicaux ; Quant au groupe des arts visuels, il est possible qu'un tel entraînement prédispose le cerveau à être plus réceptif au nouveau matériel qu'aux stimuli surappris.
Aucun changement spécifique à l'entraînement n'a été observé dans le MMN auditif, car la latence de réponse a culminé plus tôt au post-test qu'au pré-test dans tous les groupes, y compris les groupes témoins. Cela suggère que l'exposition à des stimuli lors du pré-test pourrait être suffisante pour faciliter le processus de détection des changements. Des recherches supplémentaires sont nécessaires pour examiner si une formation plus spécifique utilisant des tâches de discrimination auditive entraînerait des changements plus importants dans les réponses MMN des personnes âgées.
Formation artistique et traitement visuel
Les participants engagés dans une formation en arts visuels ont montré des différences dans le traitement cérébral précoce lié à l’attention dans les régions postérieures du cuir chevelu pariétal-occipital. Cela suggère que la formation en arts visuels peut améliorer le traitement des caractéristiques visuelles. Aucun changement de ce type dans les premières réponses visuelles n'a été observé dans le groupe témoin ou chez les participants impliqués dans le programme de formation musicale. Étant donné que le groupe des arts visuels a montré des changements dans le traitement neuronal auditif et visuel, ces résultats révèlent une propriété importante sur la nature de la plasticité cérébrale au cours du vieillissement, montrant un transfert de bénéfices entre modalités (ici de la formation aux arts visuels au traitement auditif).
Formation artistique et contrôle cognitif
Après un court programme de formation en musique ou en arts visuels de 3 mois, des changements fonctionnels cérébraux dans le contrôle inhibiteur ont été observés dans les deux groupes de formation. Bien que les deux formes de formation aient offert des améliorations similaires (c.-à-d. P3 amélioré), elles ont notamment également donné lieu à des changements spécifiques à la formation (c.-à-d. des changements différentiels en N1 et P2 pour les groupes d’art visuel par rapport aux groupes de musique). Ces résultats concordent avec les rapports antérieurs sur les modulations induites par l'entraînement dans l'activité corticale préfrontale des personnes âgées (Voss et al., 2010; Anguera et al. ., 2013).
Au niveau comportemental, le groupe de musique a amélioré sa vitesse de dénomination des couleurs (sous-test de la tâche Stroop) entre la pré- et la post-évaluation. Cet effet de l’entraînement musical pourrait refléter une amélioration de la vitesse de traitement. Le déclin de la vitesse de traitement a été associé au déclin cognitif général (Sliwinski et Buschke, 1999) et peut avoir des conséquences majeures sur la vie des personnes âgées. En effet, la vitesse de traitement joue un rôle essentiel dans les activités quotidiennes en facilitant des fonctions cognitives importantes comme l'apprentissage et la mémoire à long terme, la compréhension, la prise de décision et la planification (Ouest, 1996; Baddeley, 2012). Des études corrélationnelles antérieures ont signalé un lien entre les compétences cognitives supérieures et la musicalité (Hanna-Pladdy et MacKay, 2011; Hanna-Pladdy et Gajewski, 2012; Amer et al., 2013). Notre étude étend ces résultats antérieurs en démontrant une relation causale entre la formation musicale et la vitesse de traitement chez les personnes âgées. Cette découverte offre l'espoir d'améliorer le processus de vieillissement grâce à des stratégies d'intervention musicale visant à renforcer les compétences cognitives.
Nos résultats ont également révélé des changements dans les corrélats neuronaux du contrôle inhibiteur au cours d'une tâche GoNoGo après seulement 3 mois d'entraînement. Dans les essais NoGo, les deux groupes d'entraînement, par rapport au groupe témoin, ont montré une amplitude P3 accrue et prolongée sur l'hémisphère gauche au post-test, tandis que le groupe de musique a également montré des effets similaires sur l'hémisphère droit. Cela concorde avec une étude antérieure menée auprès de musiciens experts plus âgés (Moussard et al., 2016 ). Il est à noter que l’amélioration de NoGo-P3 dans les sites frontaux gauches a en fait augmenté la symétrie hémisphérique après l’entraînement. Cette découverte est intéressante car de tels résultats pourraient être interprétés comme un processus compensatoire du vieillissement cognitif. Des travaux de neuroimagerie ont montré que les personnes âgées très performantes recrutent des ressources supplémentaires et engagent le cortex préfrontal de manière bilatérale lors de tâches exigeantes (Cabeza, 2002; Cabeza et al. , 2002; Grady, 2012; ;Du et al., 2016). Il est possible que l’entraînement des deux groupes ait amélioré le fonctionnement des régions frontales gauches du cerveau qui avaient décliné avec l’âge. Lors des essais Go, le groupe de musique a montré une amplitude P3 améliorée après l'entraînement par rapport aux groupes d'art visuel et de contrôle. P3 peut refléter la fermeture du traitement d'inhibition d'une réponse manifeste (Gajewski et Falkenstein, 2013 ) ou l'évaluation continue d'une intention d'inhibition (Liotti et al. ., 2005). Ainsi, nous en déduisons que la formation musicale améliore les mécanismes de supervision qui agissent pour garantir les réponses souhaitées et renforcer la surveillance des interférences (Moreno et al., 2014; Moreno et Farzan, 2015).
Les effets spécifiques à l'entraînement ont été exprimés dans les ondes P2 et N2, dans lesquelles nous avons observé des effets apparemment opposés pour chaque groupe d'entraînement : les étudiants en musique ont montré une augmentation des amplitudes P2 et une diminution des amplitudes N2 après l'entraînement, tandis que le groupe des arts visuels a montré une diminution des amplitudes P2 et une augmentation des amplitudes N2. (les contrôles sans contact n’ont montré aucun changement). Dans notre étude précédente auprès de jeunes adultes suivant une instruction musicale (Moreno et al., 2014), un P2 plus grand a également été observé dans le groupe de musique. Parallèlement aux résultats actuels, cela suggère que P2 est un marqueur commun de la plasticité du contrôle inhibiteur du cerveau tout au long de la vie. On pense que la vague P2 indexe la capacité à construire une représentation du contexte actuel de la tâche et de la réponse comportementale associée dans les premières étapes du traitement (Gajewski et Falkenstein, 2013). Les preuves électrophysiologiques suggèrent que cet aspect précoce du traitement visuel est lié à des fonctions cognitives supérieures en facilitant des représentations internes plus fortes de stimuli comportementaux pertinents (Moreno et al., 2014; Moreno et Farzan, 2015). Ici, la plus grande amplitude P2 dans le groupe de musique peut refléter un traitement plus précoce et une représentation plus forte des stimuli ainsi que l'appariement de réponse (ou de non-réponse) approprié, renforcé par l'entraînement. Nos résultats ont également montré une réduction du N2 après une formation musicale, ce qui est cohérent avec des travaux antérieurs menés auprès de jeunes adultes (Moreno et al., 2014). N2 a été décrit comme un marqueur d'inhibition et de détection de conflits/charge d'attention (Nieuwenhuis et al., 2003; Burle et al., 2004; Falkenstein, 2006). Nous interprétons cet effet en conjonction avec l'augmentation de l'amplitude de P2 de telle sorte que l'entraînement musical facilite la dissociation de la planification stimulus-réponse souhaitée et non souhaitée, reflétée par une augmentation de P2, et cet effet accru de P2 réduit par la suite le besoin de processus de contrôle cognitif reflété par une diminution de N2 (Bokura et al., 2001; Nieuwenhuis et al., 2003; Donkers et van Boxtel, 2004; Kok et al., 2004).
D'un autre côté, la tendance opposée observée dans le groupe de formation en arts visuels peut suggérer que cette forme de formation a réduit les exigences perceptuelles précoces (réduction de P2). Ceci est plausible étant donné que notre GoNoGo était un paradigme visuel (même domaine que la formation en arts visuels). Les améliorations ultérieures dans le N2 ultérieur peuvent refléter une meilleure détection des conflits post-perceptuels et une meilleure inhibition de la réponse. Nos résultats concordent bien avec les preuves existantes suggérant un impact des instructions d'art visuel sur la voie visuelle ventrale (Draganski et al., 2004; Op de Beeck et Baker, 2010; Pollmann et von Cramon, 2000) ainsi que des réseaux cérébraux cognitifs d'ordre supérieur (Chamberlain et al. , 2014). Dans l’ensemble, la nature spécifique des changements induits par les instructions liées à la musique et aux arts visuels indique que ces formes divergentes d’entraînement offrent des influences uniques sur la fonction cérébrale ainsi que des améliorations des compétences générales du domaine en dehors du champ direct de l’entraînement (c’est-à-dire le contrôle inhibiteur).
Limitations
Une limite de la présente étude est que notre échantillon présentait un déséquilibre entre les sexes, ce qui pourrait affecter la généralisabilité de nos résultats. Les trois groupes comprenaient une proportion beaucoup plus grande de femmes que d’hommes. Chez les femmes âgées, la diminution du taux d’œstrogènes qui survient après la ménopause peut affecter les fonctions cognitives. Des recherches plus approfondies devraient garantir une représentation plus équilibrée des hommes et des femmes et également surveiller plus étroitement si les participantes reçoivent un traitement hormonal substitutif.
Dans la présente étude, le groupe témoin passif a été ajouté pour évaluer les effets test-retest après le test (c'est-à-dire une amélioration qui serait due à la répétition de la même batterie de tests une seconde fois). Cela nous a permis de mieux identifier la nature et la spécificité des améliorations lors du post-test. Le but de l'évaluation de suivi était de tester si ces améliorations (ou changements neuronaux) liés à l'entraînement étaient maintenues après l'arrêt de l'entraînement. Étant donné que le groupe témoin n’a reçu aucune formation, il n’était pas justifié de tester le maintien des avantages/changements liés à la formation pour ce groupe. Bien que le groupe témoin ait montré peu de changements dans ses performances et son activité cérébrale, nous ne pouvons pas exclure la possibilité que des différences soient apparues plus tard en raison de tests répétés ou d'un déclin normal lié à l'âge. Il convient cependant de noter que notre étude ne visait pas à tester directement si la formation artistique ralentissait le déclin cognitif : un suivi beaucoup plus long aurait été nécessaire à cet effet (car les personnes âgées en bonne santé ne présentent généralement pas de déclin mesurable après 3 mois). . Les futures études longitudinales devraient envisager d'ajouter des tests supplémentaires pour le(s) groupe(s) témoin(s) ainsi qu'un suivi plus long.
Enfin, bien qu'assez intensif (trois séances par semaine), le programme de formation était relativement court (3 mois). Cela pourrait avoir limité les avantages potentiels de l’entraînement musical sur les performances comportementales. De futures études pourraient envisager des périodes de formation plus longues, afin de mieux refléter l'expérience réelle de l'apprentissage de la musique.
Conclusion
Collectivement, nos résultats élargissent les connaissances actuelles sur la neuroplasticité dans le vieillissement en démontrant des changements dynamiques dans la fonction cérébrale des personnes âgées suite à une formation à court terme en musique et en arts visuels. Bien que les changements neuroplastiques restent modestes, en particulier pour les mesures comportementales, nos résultats offrent une preuve causale claire que le cerveau âgé est plus plastique qu'on ne le pensait traditionnellement et suggèrent de nouvelles possibilités d'entraînement cognitif et de réadaptation. Les études futures devraient utiliser des programmes d'entraînement plus longs pour explorer les effets de transfert dans la vie quotidienne. Avec la croissance de la population vieillissante dans la société moderne, il existe un besoin urgent de trouver des remèdes qui contrecarrent le déclin cognitif, améliorent le processus de vieillissement et, à terme, réduisent les coûts liés à la santé. Notre étude établit que les programmes d’art visuel et de musique pourraient constituer des solutions efficaces, engageantes et rentables pour stimuler la plasticité cérébrale des personnes âgées. Ces programmes pourraient améliorer les compétences essentielles, renforcer le contrôle inhibiteur et, à terme, améliorer la qualité de vie des personnes âgées.
Déclaration d'éthique
Le protocole expérimental a été approuvé par le comité de recherche humaine et d'éthique du Baycrest Centre, Toronto, ON, Canada.
Contributions d'auteur
CA et SM ont conçu les expériences. YL, AM et GB ont réalisé les expériences et collecté les données. CA, AM, GB, YL et JS ont analysé les données. CA, AM, GB, YL et SM ont interprété les résultats des expériences. CA, AM et SM ont rédigé le manuscrit. Tous les auteurs ont édité et révisé le manuscrit et ont approuvé la version finale du manuscrit.
Financement
Cette recherche a été financée par une subvention (MOP 106619) des Instituts de recherche en santé du Canada (IRSC) à CA, du ministère du Développement économique et de l'Innovation de l'Ontario à SM, au Conseil de recherches en sciences naturelles et en génie (RGPIN-06196-2014 à SM) et l'Institut national sur la surdité et autres troubles de la communication du NIH (R01DC016267 à GB).
Déclaration de conflit d'intérêts
Les auteurs déclarent que la recherche a été menée en l'absence de toute relation commerciale ou financière pouvant être interprétée comme un potentiel conflit d'intérêts..
Remerciements
Nous tenons à remercier nos professeurs de musique et d'arts visuels, Donna Takacs et du Royal Conservatory of Music, Madeline Chan, Courtney Smith, Buddhika Bellana, Tristan Watson, ainsi que tous les participants à notre étude.